�X�̐��I���n�܂�܂����@(��30��)
�����@�����x����_�C�i���@�i9�j�@����
���{���؎�q�����������E���k��w���_�����@�@�]�h�m�i
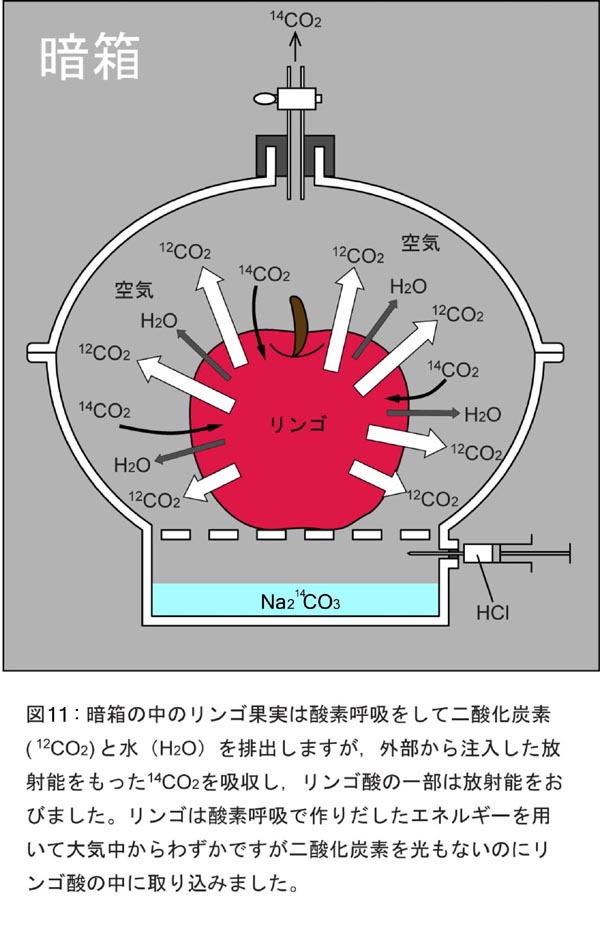
�@�A���̖����x�������̃_�C�i���́A�����Ƃ͈���ėL�@������葱���邱�Ƃʼn��ȊO�ɕ��@�͂���܂���ł���(��26��)�B�A�������L�@���ƂȂ�ƒ��[�����g�D��`���w(��16��A�}7)�ł̍זE����Ƃ��̌�̍זE�����i�����A�j�_��^���p�N���Ȃǂ̐������j�A�����Ă�������łɂ��邽�߂̍זE�ǁi�Z�����[�Y���\�Ƃ��鑽���ނ�O�j���Ȃǁj�������킸��葱���邱�Ƃł����B�����̍�Ƃ��i��@�\�I�ȕ����i�y�f�A��y�f�Ȃǁj�̐��Y�̏o���_�̓A�~�m�_�ł���A�����͒Y�f���i�ɖ��@�̒��f���f���A�~�m��Ƃ��Č��ѕt���邱�ƂŎn�܂�܂���(��28��A�}18)�B�Ƃ���ŁA�A���̐���̗L��l�����߂Ă����͍̂��n�ł������i��16��j�A���Ɩ����ɐ��炷�鍪�n�͒n�㕔����ڑ�����ė��铜�������ɗ������_�C�i�����ғ������A�����Ă���̂ł��傤���B�L�@�����Y���ێ����Ȃ������~����^���ɂ����_�C�i����n���ł������������邽�߂ɂ́A������Y�f���i�Y����H�v���K�v�ł��B�܂�A���n�͎���Èł̒��œ�_���Y�f(CO2)���Ҍ��Œ肹����Ȃ��̂ł����A����͓���Ȃ��Ƃł͂Ȃ��A�����S�����s���Ă���A���ɔ��������ʓI�d�g�݂ł����i��20��A�}11�j�B�����ō���́A�悸�����S�͂ǂ̂悤�ɂ���CO2����荞��ł����̂��w�Ԃ��Ƃ���n�߂܂��傤�B�q���g�́A�������ɍۂ���CO2���Ҍ������͈̂Ô����ƌ�����ATP��NADPH�Ɉˑ������y�f�����ł��B�Ȃ�Βn���ł��A�Ô������i�ޏ������������Ă���CO2���Ҍ��Œ肵�ĒY�f���i�������o����͂��ł��B�����ɐA���������I�����̃_�C�i������]����������̍H�v������܂��B
���n���̓����v�ƘA�g���čs���Y�f���i�����V�X�e���������S�Ɋw��
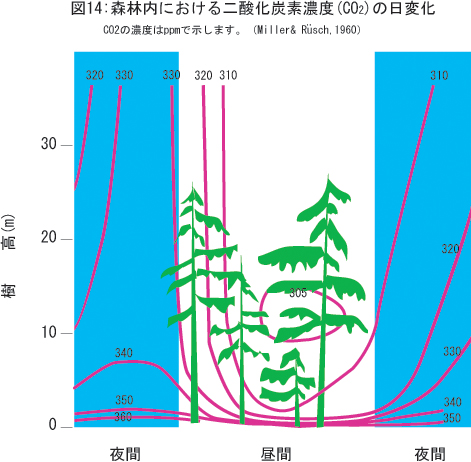
�@����܂Œn���Ƃ����Èł̔��������ł̍��n�̐����ƍ����ϓ���(��21��A�}13)�A�n�㕔�ł̏A���^���ɑΉ���(��19��)�A�������ɕ��ՓI�ȑ̓����v�̑��݂��������܂����B���̍��n�͐X�ђ��̒n�\�ʂł̍���CO2�Z�x(��23��A�}14)�����I�݂Ɏ���̔���Ɋ������Ă����̂ł��B�������ĐX�т̍��n�̔��B�ɍD�s���Ȋ������������炱���A�h�Ђɋ������n�l�b�g���[�N��n�w�\�w�Ɉ�āA���R�ЊQ�����{�̎��R����邱�Ƃɂ���^�ł���̂ł��B���B�͂��������X�̎��R�ȉc�݂ɂ��A�X���L����{���I�ȈӋ`��F�߂�ׂ��ł���A����͎��̍��n���X�̒��ł����ɏ�v�Ɉ���Ɋ|�����Ă��܂��B
�@
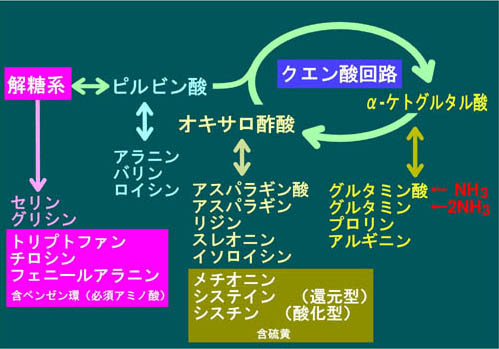
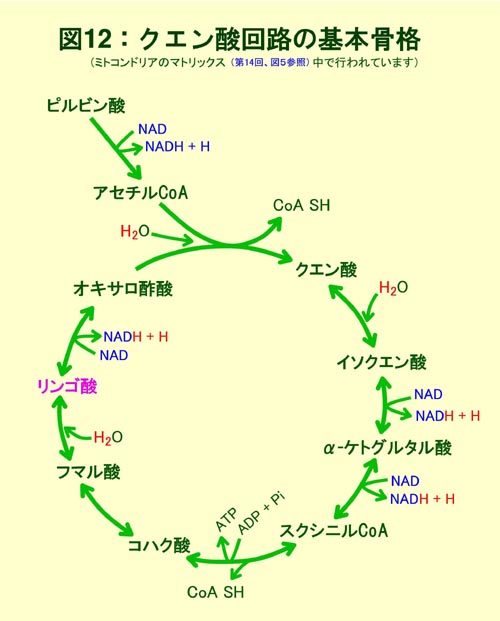
������23���ŏq�ׂ��悤��CO2�͓����זE��������ɂ��s���ŁA�זE�̔|�{�ɂ͑�C���̂��̔Z�x�����߂邱�Ƃ��K�v�Əq�ׂ܂����B���̓_���炷��ƁA�A�������ɔ�������V�X�e���Ƃ͌����܂��A�A�����_�C�i����]�ɕK�v�ȑS�ẴA�~�m�_�Q�i�}17�j�𑵂��邱�Ƃ́A�Y��������ӂ̖{���A�n����N�G���_��H�i��18��A�}10����20��A�}12�j��ATP/ADP��Œ��߂�����x�ł͕s�\�ł��B���̒��x�ł��x���[����L����K�{�A�~�m�_�Q�i��28��A�}17�j�Ȃǂ������ł��܂����A���̂��d�v�ȃA�~�m�_�̌`���p�̒Y�f���i�̋����͏o���܂���B�ŏd�v�ȗ������܂ރA�~�m�_�Q(��28��A�}17)�̓I�L�T���|�_�R���̃A�X�p���M���_�Q�̈���ł����A�N�G���_��H���̂̓I�L�T���|�_�ƃA�Z�`��COA�Ɛ��Ƃ��N�G���_�k���y�f�ɂ���ĕs�t�I�ɍ����]���n�߂���̂����ɁA��H�����烿�|�P�g�O���^���_��I�L�T���|�_���A�~�m�_���i�ɂ��邽�߂Ɉ����������Ƃ̓N�G���_��H���̂��~�߁A���̃_�C�i�����̂̉ғ����~�߂鎖�ɂȂ��Ă��܂��܂��B���������Ċe��@�\�I�A�~�m�_�Q�̌`���̂��߂̒Y�f���i������S�����ƂɂȂ�̂��A�}20�Ɏ����������S���s�Ȃ��Ă����Y��������ӂ̃o�C�p�X�ł��B�}10���̉n�̍Ō�ɗ���z�X�z�G�m�[���s���r���_�iPEP�j��PEP�J���{�L�V�L�i�[�[�Ƃ����y�f�������āAPEP+CO2+ADP�̃I�L�T���|�_�{ATP�̔�����CO2���ԂɊҌ��Œ肵�Ă��܂��̂ł��B���̌��ʁA�ŏI�I�ɂ̓����S�_�ƃA�X�p���M���_���ɗ��ߍ��ނ悤�ɓ����܂��B���̌��ʁA�I�L�T���|�_�ɋN������A�X�p���M���_�R���̊ܗ����̃A�~�m�_�`���̐��𐮂��邱�Ƃ��\�ƂȂ�܂��B�n���̈ÈłŎ��͂̍���CO2�Z�x���I�݂ɒY�f���i�Ƃ��Ď�X�̋@�\�I�����q�������A��y�f�i�r�^�~���j��y�f�^���p�N���̌`�����\�Ƃ����̂ł��B�@
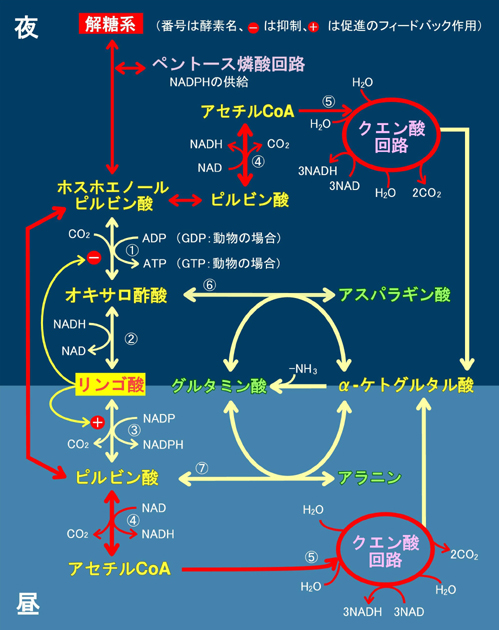
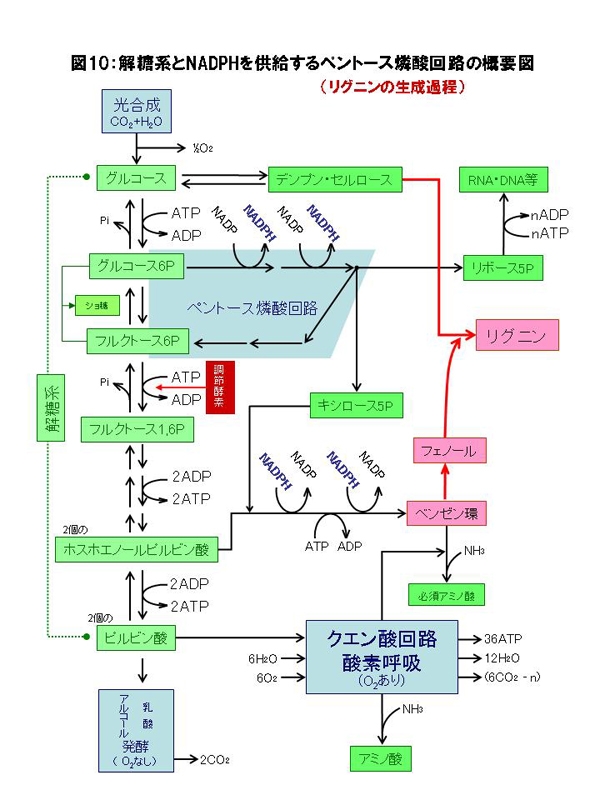
�}20�F�A���ɂ�����Y�f���i��[�̂��߂̒Y��������ӂ̃o�C�p�X�Ƒ̓����v
�@PEP�J���{�L�V�L�i�[�[�@�A�����S�_�E���f�y�f�@�B�����S�_�y�f�@�C�s���r���_�E���f�y�f�@�D�N�G���_�k���y�f�i�s�t�I�����j�@�E�O���^�~���_-�I�L�T���|�_�A�~�m��]�ڍy�f�@�F�O���^�~���_-�s���r���_�A�~�m��]�ڍy�f |
�@�}20�������悤�ɂ����ɃA�~�m�����Ă���̂̓N�G���_��H�R���̃��|�P�g�O���^���_������ƂȂ�O���^�~���_����̃A�~�m��̓����Ƃ��̈ڂ��ւ��ł��B�A����PEP�J���{�L�V�L�i�[�[�͓����Ƃ͈���āA��y�f�Ƃ���GDP�ł͂Ȃ�ADP���W���ATP�Y���܂����A�~�g�R���h���A�ɂ��������̃_�C�i���̉ғ����ێ����邽�߂ɂ��A���`������ADP���������Ȃ���ɂ߂ďd�v�Ȗ�����S���A�X�p���M���_�i�ܗ����A�~�m�_��ADP���̒��ԃv�����������̐��Y�ɕK�{�j���������A�~�g�R���h���A���̃N�G���_��H�ő�ʂɋ�������ė���NADH���_�C�i����]�Ɏg�킸�ɁANADH�ɒ~�����Ă���d�q�������S�_�Ɉڂ������鎖�ŁA�A���̍��n�͖�Ԃ̊ԂɎ�X�̗L�@�����Y�̂��߂̑f�ޒ����łȂ��A������p���Ă̎��ۂ̗L�@�����Y�������������Ă����̂ł��B�ߏ�ɒ~�ς��������S�_�͑O�i�K�̍y�f�������t�B�[�h�o�b�N�}�����i���H�̌�ʏa���l����ƕ���܂��ˁj�A���̏�Ɍ��`�����ł�ATP/ADP��̑�����}10���̒��ߍy�f�̍쓮���y���g�[�X�ӎ_��H�ғ��ɍD�s���ȏ����ł��̂ŁA���`�����ɂ͊e��̒Y�f���i���������łȂ��A�L�x��ATP,NADPH�����Ɖ]���L�@�����Y�ɍœK�Ȋ��ގ��ɂȂ����̂ł��B���̎��͗L�@�����Y���������邱�ƈȊO�����̃_�C�i������]���������i�̖��������A���ɂƂ��Ă͂ƂĂ��厖�Ȃ��Ƃł��B
���n�l�b�g���[�N����Ă���̒m�b
�@�Ƃ���ŁA�}20��������x�������܂��傤�B�̓����v�̖�ԂɑΉ����ċN�����Ă��������́A�n�㕔����ڑ�����ė����������n��ʂ��đ�ӕ������A�n���̈Èł�PEP�J���{�L�V�L�i�[�[�������Ă�CO2�̊Ҍ��Œ�ł�(�}20)�BCO2�͍ŏI�I�ɂ̓����S�_�Ƃ��Ē~���������ŁA�r������I�L�T���|�_�̓A�~�m�_�Ɏp��ς��čs���܂��B���̊Ԃ̔����̓N�G���_��H�Ƃ͈���Ĉꌩ�t���ƂȂ�܂����A���̂��Ƃ͌�������������S�_�ͤ��Ԃɂ��ғ���������~�g�R���h���A���甭������NADH�����̃_�C�i���̉ғ������ł͏���ꂸ�ɗ]��NADH�Ȃ�d�q��~���钙���ɂ̖������ʂ����Ă����̂ł��B���ۂɂ�CO2�Œ�͌��`�����Ői�s���锽���ŁA�N�G���_��H���t�����Ă����ł͂���܂��A���̎����ɂ̓N�G���_��H�ւ̗L�@�_����������܂��̂ŁA�_�f�����݂��Ă��_�f�ċz��ATP���ʂɐ��Y����K�v���ቺ���A��Ԃɂ͒n�㕔�����܂苁�߂Ȃ�������ATP���ʂɏ���đ�n���狭���ɏW�߂Ēn�㕔�ɑ�������傫�ȍ�����K�v�Ƃ��Ȃ����ԑт������̂ł��B���ꂪ�}13�̉t�샊�Y���̒J�ƂȂ��Č���A�l���m�L�Ȃ�c���͒ቺ���ď��t����Ă��܂��܂��B���ꂪ�S�����ɋ��ʂ̑̓����v�̕\���Ƃ��Ă̏A���^���Ȃ̂ł��B�����ɉߏ�ɗ��܂��������S�_��PEP�J���{�L�V�L�i�[�[�ɑ��Ă͕��̃t�B�[�h�o�b�N�ɓ�����CO2�̌Œ葬�x�����������n�߁A�����ւ̏������n�܂�܂��B
�@
�������Ē������ꂽ�����S�_���ɒ�����ꂽ�d�q�͑̓����v�̈ꗃ��S����������n�܂钋�Ԃɑ������鎞���ɈÈł̒n���ł������͂����A�n�㕔�����߂邾���̑�ʂ̐����������ł��邱�ƂɂȂ�܂��B����̓����S�_�������S�_�y�f�ɐ��̃t�B�[�h�o�b�N�ɓ����ăs���r���_���������n�߁A�N�G���_��H�̉�]�𑣂����܂��i�}20�j�B�₪�đ�ʂ�ATP�̐��Y���n�܂�A��n���琅���������W�ߋz�����Ēn�㕔�ɉ^�яグ��傫�ȍ����ނ��ƂɂȂ�܂��B���������}13�̉t�僊�Y���̎R���o�����A�t�͖ڊo�߂ēW�J�i�c������j���n�߂܂��B�A���t�̏A���^���́A�n���ɐ����鍪�n�ł̑̓����v�̖{�̂̓����S�_�̒~�ςƏ���Ƃ��W����Ӑ��䂾�����̂ł��B�����A���̎����ɂ͍��n�ł��N�G���_��H�̉����ȉғ����n�܂�̂ŁA�A�~�m����̓�����ƂȂ郿�\�P�g�O���^���_�̕�[�������͉��P����ăA�~�m��]�ڍy�f�������n�ߊe��̃A�~�m�_�̐��������\�ɂȂ�܂��B����ɂ��_�C�i���ғ��Ŏn�܂�ATP/ADP��̏㏸���傢�ɍv�����܂��B�Ɠ����ɁA�����S�_�ƃs���r���_�Ƃ̊Ԃ�G�}����y�f�A�����S�_�y�f�͕�y�f�Ƃ���NADP���g���̂ŁA�Èł̒n���ł͕s������NADPH���A�y���g�[�X�ӎ_��H�ȊO������⋋���邱�Ƃɂ��Ȃ�A�L�@�����Y�̐��̐����ɂ��D�s���ŁA�X�����̃_�C�i���̉�]�����x����傫�ȏ����ƂȂ�܂��B
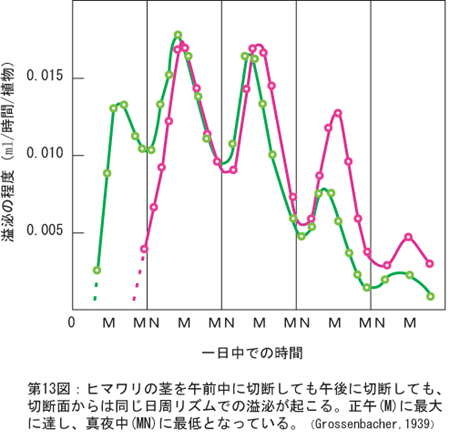
�@��19���ŁA�����l���m�L���ӂ��҂̂悤�ɑ��z���R�A�ɒ��ޑO���疰���Ă��܂��ʐ^�����������܂������A�z������Ƀ}���Ȃł��邽�߂ɍ��ɋ������Ă��鍪���ۂɗ^����ɏ[���ȓ������������Œ~���I�����Ȃ�A�������t����ď��U�𒆎~���邱�Ƃ̕����������̂ł��傤�B�t���J���Ă����ɂ́A�����ێ���ATP������˂Ȃ炸�A�n�����̔��B�Ɉڑ�����ė��铜����L���Ɋ��p�����Ȃ������ɈႢ����܂���B������CO2�𑽗l�ȒY�f���i�`���ɗ��p���邽�߂ɖ���ɏA�������������̂ł��傤�B���̎��ɂ̓����S�_�����܂�n�߂܂����A���̗ʂ��������CO2�Œ���~�܂�A�t�Ƀ����S�_���̂������S�_�y�f�������������܂��̂ŁA�s���r���_�\�A�Z�`��COA�\�N�G���_��H�̖{�����o�C�p�X������⊮���邱�ƂɂȂ��ē�d�ɑ������邱�ƂɂȂ�A���ʂ�ATP���l�����āA�G�l���M�[�ˑ��̋z���E��������ɏ[���ȍ����āA��������Ăя��t���J�����ɈႢ����܂���B�F����̓��������A�����l���m�L�̍��n���ł̕����̓��Ԃ��w�I�ɉ�͂��āA�̓����v�̖{�̂������ė~�������̂ł��B
|



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}